|
 |
|
3−1.落葉林から常緑林への交代
|
港大橋、中島大橋、玉津環境センターのどの花粉ダイアグラムをみても、7,000年代にかかるとナラ類は急速に減少しはじめ、それに代ってカシ類が急増する。そして6,000年前には、両者は完全に入れかわっている。それにともなってブナをはじめ、ナラ類とともに森林をつくっていたトチノキやシナノキ、カバノキの仲間なども北国へ、あるいは高い山地へ移動し、大阪湾沿岸からは姿を消してしまう。この大規模な森林の移動をナラ類とカシ類とにしぼって追跡してみょう。
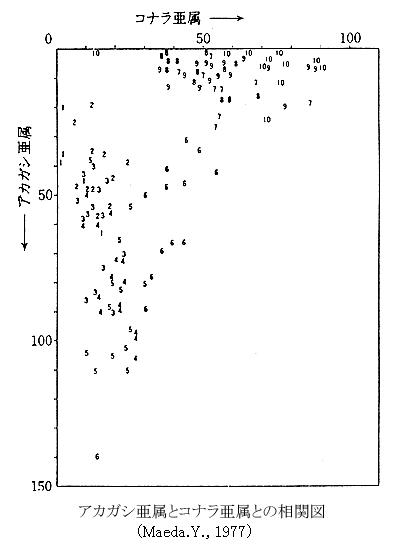 左の図は、ナラ類とカシ類の花粉化石の出現の推移を示す相関図である。この図の読み方は少し説明がいるかも知れない。いま、中島大橋のマイナス17.3メートルで採集した粘土から花粉化石を抽出分離し、顕微鏡下で検鏡するプレパラートができた。この粘土の14C年代は6,150年前である。花粉化石を種類別に読みとったところ、ナラ類が36粒、カシ類が69粒数えられた。つぎに相関図のナラ類の花粉粒を示す横軸上に36の位置をとり、いっぽう縦軸にはカシ類の69をおさえ、両者の交点に6を記入する。これは6,000年代をあらわしている。このような方法で、阪急梅田駅の地下、港大橋、中島大橋で分析した花粉化石の数をプロットしたのが左の図である。 左の図は、ナラ類とカシ類の花粉化石の出現の推移を示す相関図である。この図の読み方は少し説明がいるかも知れない。いま、中島大橋のマイナス17.3メートルで採集した粘土から花粉化石を抽出分離し、顕微鏡下で検鏡するプレパラートができた。この粘土の14C年代は6,150年前である。花粉化石を種類別に読みとったところ、ナラ類が36粒、カシ類が69粒数えられた。つぎに相関図のナラ類の花粉粒を示す横軸上に36の位置をとり、いっぽう縦軸にはカシ類の69をおさえ、両者の交点に6を記入する。これは6,000年代をあらわしている。このような方法で、阪急梅田駅の地下、港大橋、中島大橋で分析した花粉化石の数をプロットしたのが左の図である。
こうして作製したナラ類とカシ類の相関図をみると10,000年前から7,000年前まではナラ類が圧倒的に多く、5,000年前以降ではカシ類が多くなり、両者の関係が逆の相関にあることが明瞭に示されている。さらに6,000年代は両者の関係が入れかわった年代であることも見事に表現されている。全体としてみると、年代を示す数は原点に対して直交する方向に並び負の相関関係のパターンである。このことは、ナラ類とカシ類の生存する気候環境が異なることも意味している。もし、同じような環境に生育する植物どうしであれば、正の相関のパターンである原点からのびる対角線に年代が集中するはずである。
|
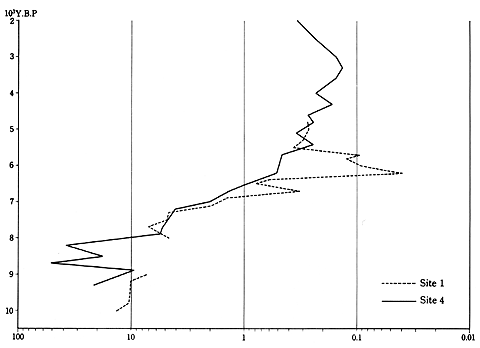
コナラ亜属とアカガシ亜属の花粉化石の出現比
site1は港大橋、site4は中島大橋 (Maeda.Y., 1977) |
ナラ類からカシ類へ移る過程をもうひとつの方法でチェックしてみた。それは両者の消長のもようを比の関係で検討したことである。さきの相関図の読み方で例示した中島大橋のマイナス17.3メートルの6,150年前の試料で説明すると、ナラ類の花粉粒36をカシ類の花粉粒69で割れば、0.522になる。これは、この当時、ナラ類の花粉はカシ類の花粉粒の約半分しかなかったことをあらわしている。花粉粒の多少が、そのまま当時の森林の分布(被度)につながるとは思わないが、ひとつの傾向は示していると考えてよい。このような方法でナラ類とカシ類の花粉化石の出現比を求めて図化したのが83ページの図である。これによれば10,000年前から8,000年前にかけては、カシ類の数十倍も出現していたナラ類の花粉化石が7,000年前ごろから両者は1対1になり、6,000年前にはついにカシ類が10倍近い数で出現していることがはっきりあらわれている。
このような7,000〜6,000年前にかけての大規模かつ急速な森林交代は、大阪湾沿岸以外でも進行していたのか。74ページの明石川の玉津環境センターでの出現傾向をみても、大阪湾沿岸同様に、両森林の交代がじつに見事にキャッチされている。しかもここではアカホヤ火山灰がちょうどその時期にあたっている。アカホヤ火山灰の14C年代は約6,400年前である。
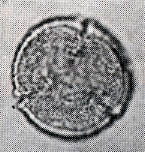
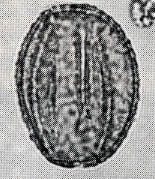
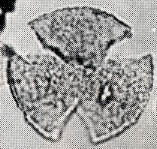
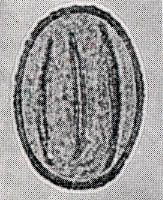
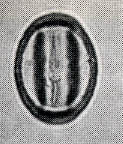
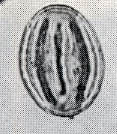
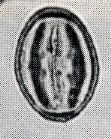
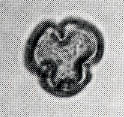
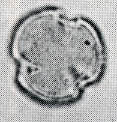
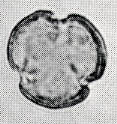
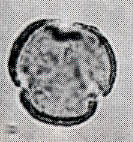
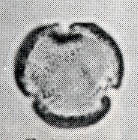
花粉化石は前にも述べたように肉眼では見えないほどの小さいものである。ナラ類は30ミクロン前後で、カシ類は25ミクロン前後である。1ミクロンは1,000分の1ミリであるから、肉眼で見分けられないのはとうぜんである。顕微鏡で数百倍(400倍程度)に拡大して種類を識別するのであるが、その判定には問題はないだろうか。普通の光学顕微鏡下ではナラ類の花粉の表面にはやや顕著な粒状突起がみとめられ、それに対してカシ類は一般にナラ類よりも小さく、その表面にも突起はなく、比較的平滑である。私はこの両者の違いをさらに明確にすべく、松岡数充さん(長崎大学)と共同で日本産のコナラ属の表面形態を、走査型電子顕微鏡を使って調べた。
|
 |
 |
 |
 |
| 1a |
1b |
2a |
2b |
 |
 |
 |
 |
| 3a |
3b |
4a |
4b |
 |
 |
 |
 |
| 5a |
5b |
6a |
6b |
|
|
|
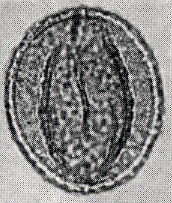
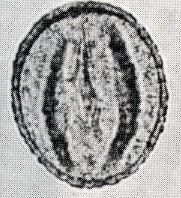
カシ類とナラ類の光学顕微鏡写真 原著:約800倍 (大阪湾累層)
1〜6.カシ類 (アカガシ亜属) 7〜11.ナラ類 (コナラ亜属)
(Maeda.Y., 1977)
|
(次ページへ続く)
|
|